|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Le chapitre
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
Dernière mise à jour le
|
![]() I. Recherche de parenté chez les vertébrés avec établissement de phylogénies
I. Recherche de parenté chez les vertébrés avec établissement de phylogénies
|
• T.P. 1
|
|
A. Parenté anatomique
Belin, doc. 1B, p. 21 : le membre antérieur des vertébrés1. Notion d’analogie et d'homologie
Documents : l’aile qui permet de voler et le plan d'organisation du membre antérieur des Vertébrés (Belin, doc. 1B, p. 21)
- Les ailes de la chauve-souris et de la mouche ont une même fonction (prendre appui sur l'air) assurée par une membrane de grande surface soutenue par une armature. Mais la structure des organes qui constituent l’aile est très différente. Cette ressemblance constitue une analogie.
- Les membres des Vertébrés présente un même plan d'organisation en 3 segments, chaque segment comportant une même composition osseuse. Avec un même plan d'organisation, les membres des Vertébrés présentent des adaptations. On parle alors de structures homologues pouvant être héritées d'un ancêtre commun (Définition Belin, p. 30).
- Comparaison du plan de construction des membres antérieurs de Vertébrés

2. Notion de caractères dérivés
Belin A1 et 2, p. 22
- État ancestral ou primitif
État dérivé
3. Bilan
- Dans un groupe d'espèces, une espèce A est plus étroitement apparentée à une espèce B qu'à toutes les autres, si elle possède avec l'espèce B un ancêtre commun exclusif (que les deux espèces A et B ne partagent pas avec les autres espèces). Les ancêtres communs se trouvent toujours aux nœuds de l'arbre.
- Un état d'un caractère qui résulte d'une innovation génétique est dit dérivé (on dit aussi caractère dérivé). Les relations de parenté sont établies en prenant uniquement en compte les caractères dérivés.
- La notion de caractère dérivé est cependant relative. Elle est définie par rapport au groupe d'espèces dont on recherche les liens de parenté. Ainsi, au sein des Vertébrés, la possession de doigts est un caractère dérivé mais au sein des tétrapodes - qui possèdent tous des doigts - c'est un état ancestral (qui ne peut servir pour établir des degrés de parenté au sein des tétrapodes).
- Un fossile n'est jamais un ancêtre commun. Un ancêtre commun est toujours hypothétique, défini par toutes les innovations (caractères dérivés) des espèces qui en dérivent et possédant également les caractères ancestraux des groupes dans lesquels il est inclus.
B. Parenté embryologique
1. À partir de critères comme présence d’amnios
Voir fiche TP. 1 et un autre document : structure de la mandibule chez les Vertébrés, Belin, doc. A, p. 20.
2. À partir de similitudes de développements dans les premiers stades
Voir fiche TP. 1.
C. Parenté moléculaire
1. Notion de molécules homologues
- Certaines molécules assurent une même fonction chez un grand nombre d’êtres vivants, elles sont constituées d’une séquence d’acides aminés possédant une forme tridimensionnelle spécifique permettant d’assurer la fonction. C'est le cas du cytochrome C qui intervient dans la respiration cellulaire de tous les organismes qui respirent, des globines alpha et bêta qui constituent l’hémoglobine des êtres vivants à sang rouge ou de la myoglobine qui assure le stockage du dioxygène dans le muscle.
- Ces molécules diffèrent d’une espèce à l'autre par la séquence d’acides aminés tout en conservant l'aspect tridimensionnel caractéristique de la fonction.
- Pour apprécier le degré de parenté entre 2 espèces, on mesure d’écart qui existe en recensant les différences qui existent entre les séquences d’acides aminés de la molécule.
2. Notion de distance moléculaire pour construire un cladogramme
Belin, doc. B, p. 27.
D. Construire un arbre phylogénétique en se fondant sur le partage de caractères dérivés
Des synonymes - Arbre phylogénétique = Arbre phylogénique = Cladogramme1. Classifications traditionnelle et phylogénétique
Belin, A1, p. 26.2. Le principe de construction d'un arbre phylogénétique
- La parenté
- La parenté est chercher le groupe frère (et non l’ancêtre) avec lequel il partage un caractère exclusif. Cela conduit à déduire certains des caractères que devait posséder un ancêtre hypothétique, l’ancêtre au sens propre restant toujours inconnaissable.
- Phylogénie et cladogramme (arbre phylogénétique)
- Une phylogénie se traduit par une figure arborescente (le cladogramme) fondée sur le concept de descendance avec modification des caractères après avoir reconnu l'état primitif et l'état dérivé d'un caractère.
Le cladogramme traduit les liens de parenté, les nœuds sont fondés sur les caractères dérivés exclusifs. Le cladogramme dit « qui est le plus proche parent de qui » et « non qui descend de qui ».
- Une phylogénie se traduit par une figure arborescente (le cladogramme) fondée sur le concept de descendance avec modification des caractères après avoir reconnu l'état primitif et l'état dérivé d'un caractère.
3. Une approche de la construction de l'arbre. phylogénétique des Vertébrés.
Belin, doc. 2B, p. 23 : Truite, Homme, Pigeon et Chauve-souris.
- Travaux pratiques : recherche de critères qui permettent de définir une innovation à partir d'un caractère primitif et d'un caractère dérivé (utilise Phylogène). Se reporter aux fiches TP. 1 et TP. 2.
- Définition.
- Taxon : groupe d'organismes formant une unité bien délimitée à chacun des niveaux hiérarchiques de la classification : espèces, genres, familles, les ordres.
- Choane : narine interne.
- Le principe des arbres phylogénétiques : le partage de caractères dérivés définit l’appartenance à un groupe.
E. Une espèce fossile ne peut pas être considérée comme ancêtre d'un groupe.
Se reporter à la fiche TP. 1 et au corrigé. Belin, Unité 3, doc. A et B4-5, pages 24-25 + Fiche Place de l’Archéoptéryx dans la classification.
- Possibilité de travailler avec Phylogène (si on a le temps ou à la maison).
- Protocole identique à celui qui a été décrit au TP utilisant Phylogene, 3e étape.
- Choisir les espèces suivantes : Aigle, Archéoptéryx, Compsognathus, Crocodile, Lézard, Otarie (taxon externe) et Pigeon.
- Sélectionner les critères : bréchet, doigts (observer sur les dessins leur disposition), fenêtre mandibulaire, fenêtre temporales, plumes et queue.
F. Conclusion
De nombreux critères sont nécessaires à la classification phylogénétique des êtres vivants. C'est le recoupement de ces données comparées aux données paléontologiques et aux datations d’apparition des ordres et des espèces qui a permis la mise en place de la nouvelle classification phylogénétique. Contrairement à la classification traditionnelle qui s’appuyait sur l’anatomie et la morphologie, la phylogénie est le résultat des apports de l’embryologie, de la biologie moléculaire qui permet une nouvelle approche par la mesure des degrés de parenté.
![]() II. La lignée humaine
II. La lignée humaine
A. Place de l’Homme dans le Règne animal et parmi les vertébrés
1. Dans le Règne animal et parmi les vertébrés
2. Chez les primates
Belin p. 39 (Bonobos, Chimpanzés nains)
|
• T.P. 2
|
|
B. Parenté au sein des Primates
• Les caractères dérivés propres aux Primates : pouce opposable, les ongles remplacent les griffes, les orbites migrent vers l'avant favorisant une vision binoculaire,…
• Les caractères dérivés propres aux Haplorrhiniens : le rhinarium disparait et est remplacé par le nez, les vibrisses (moustaches disparaissent,…
• Les caractères dérivés propres aux Simiiformes : fermeture postérieure de l'orbite par une paroi osseuse,…
• Les caractères dérivés propres aux Catarrhiniens : narines ouvertes vers le bas, séparées par une fine cloison, rapprochées, …
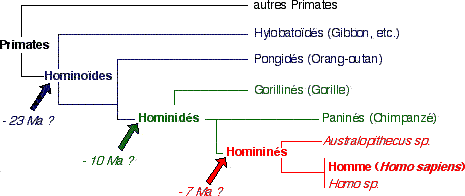 |
• Hominoïdes : super-famille regroupant les Primates dépourvus de queue.
• Hominidés : famille regroupant l'Homme, ses ancêtres et leurs collatéraux. Inclus aussi les Chimpanzés et les Gorilles (programme officiel). • Homininés : sous-famille des Hominidés, comprenant les genres Homo et Australopithecus. Ma = Million d’années |
- L'arbre ci-dessus est celui proposé pour l'épreuve du baccalauréat. D'autres arbres existent et sont produits par des méthodes différentes : http://www.snv.jussieu.fr/vie/bib/dos-doc/1documents.htm
1. Données chromosomiques
Belin, p. 39.
- 2 Documents :
- Caryotypes comparés de l'Homme et du Chimpanzé
- Phylogénie des primates à partir des réarrangements chromosomiques
- Caryotypes comparés de l'Homme et du Chimpanzé
- La comparaison entre Chimpanzé et Homme montre :
- 13 chromosomes identiques (3, 6, 7, 8, 10, 11, 14, 16, 19, 20, 21, 22 et XY) ;
- 10 chromosomes présentant des différences (qui n'ont pas de sens évolutif) : délétion, fusion (chromosome 2 humain), insertions, nombreuses inversions.
2. Données moléculaires
Des exemples ont été vus dans le T.P. utilisant Phylogène pour comparer les données moléculaires.
C. Homme et Chimpanzé partagent un ancêtre commun
Fiche documentaire ![]()
D. Critères d'appartenance à la lignée humaine
|
• T.P. 3
|
|
DE TOUMAÏ À SAPIENS : la ruée vers l'Homme - Un dossier de la Cité des Sciences de la Villette - Lien
1. Bipédie permanente.
2. Squelette de la tête et développement du cerveau
- 2 Fiches documentaires (voir TP 3) : Evolution des boîtes crâniennes et Comparaison des boîtes crâniennes et des moulages endocrâniens.
3. Activités culturelles et industrielles
Belin 2 et 3 p. 46 et 3, p. 41.4. Comment classer un fossile dans la lignée humaine ?
- Les caractères dérivés qui caractérisent la lignée humaine ne sont donc pas uniquement d'ordre anatomique, chromosomique ou moléculaire.
- La fabrication, la conservation et l'usage répétitif d'un même outil, l'acquisition d'une pensée abstraite comportant des activités artistiques et des croyances religieuses sont des marqueurs de la lignée humaine.
![]() III. Caractère buissonnant de la lignée humaine et origines d’Homo sapiens
III. Caractère buissonnant de la lignée humaine et origines d’Homo sapiens
A. Les Australopithèques
1. Premiers représentants des Homininés
Empreinte de Laetoli (Tanzanie)
Belin p. 54-55.
- Les restes fossiles et les traces des australopithèques comme Lucy nous donnent les preuves d’acquisition de la bipédie.
- D'autres données font penser que cette acquisition s'est faite sans doute par mutation d'un gène du développement.
- Ce type de mutation présente la particularité d'entraîner des modifications très importantes du phénotype pour une modification moindre du génotype.
2. La mosaïque des Homininés
Réaliser un diagramme pour représenter en parallèle la présence des différentes espèces, prévoir Orrorin (sera complétée au fur et à mesure avec le genre Homo) donc ordonnées de - 6 Ma à 0 Ma.
- Australopithèques
- Datés de 4,5 à 2,2 Ma, d’une taille de 1m à 1,30m, d'un volume cérébral de 4 à 500 cm3, bipèdes, arboricoles.
- Paranthropes (ex Australopithèques Robustes)
- Datés de 2,6 à 1,2 Ma, d’une taille de 1,1m à 1,30m, d'un volume cérébral de 5 à 600 cm3, incomplètement bipèdes, arboricoles.
- Australopithèques de l'Ouest (Abel)
- Datés de 3 à 3,5 Ma.
- Orrorin tugenensis (« Ancêtre du Millénaire »)
- Datés de 6 Ma, bipède forestier du Tchad, place l’apparition de la bipédie 2 Ma d’années avant ce qu'il était habituel de penser (4,2 Ma) et remet peut-être en cause la théorie selon laquelle la bipédie serait apparue du fait d’une évolution de l'environnement en savane suite à la surrection de la région du rift est-africain.
- L’East side story
Belin, p. 55- D'après Yves Coppens, les homininés sont apparus dans des milieux de savane arborée en Afrique de l'Est. Leur séparation avec les grands singes, qui sont tous forestiers, est datée de plus de 7 millions d’années. Leur apparition est donc synchrone de la mise en place du rift africain, qui s'est accompagnée de la régression vers l'Ouest de la forêt, habitat des grands singes, et de l'apparition à l'Est de hauts plateaux au climat plus sec favorable à la savane. Il existe donc un lien entre l'émergence des homininés et la genèse du rift : ce dernier a créé un environnement plus favorable à des primates bipèdes et il a séparé leurs populations naissantes de celles des grands singes, avec lesquels ils partagent un ancêtre commun.
B. Émergence du genre Homo au sein des Australopithèques : Homo habilis
1. Homo habilis
Datés de 2,45 à 1,55 Ma, de petite taille (1,25m), d'un plus grand volume cérébral (700 cm3), bipèdes permanents, fabriquent des outils (galets aménagés).
2. Communauté probable des Homo et des Paranthropes
Compléter le diagramme des Australopithèques.
C. Homo erectus : les premiers voyageurs qui colonisent l'Afrique et l’Eurasie
L’adolescent de Turkana
Datés de 1,8 à 0,15 Ma (40 000 ans en Asie), de grande taille (1,8m), d'un grand volume cérébral (800 à 1200 cm3), bipèdes permanents, fabriquent des outils (bifaces) et maîtrisent le feu, participent à plusieurs vagues migratoires vers l'Afrique, l'Asie (Homme de Pékin) et l'Europe (Homme de Tautavel).
D. « Néandertal » et « Cro-Magnon », deux espèces séparées il y a 400 000 ans
Belin unités 4 et 5.
- Les tests ADN mitochondrial ont permis de montrer des différences qui montrent une possible séparation ces deux espèces du genre Homo, il y a 400 000 ans.
1. Homo sapiens neandertalis
- Datés de 110 000 à 30 000 ans, de grande taille (1,7m), d'un grand volume cérébral (1200 à 1700 cm3), bipèdes permanents, maîtrisent le feu, fabriquent des outils diversifiés et respectent des rites (rites funéraires), présents en Eurasie.
- La place de l'homme de Neandertal dans l'histoire des homininés peut constituer le support d'un questionnement sur les fondements de la notion d'espèce en paléontologie, en l'absence de possibilité de tester l'interfécondité (il faut toutefois avoir donné au préalable une définition biologique de l'espèce). II est intéressant de mentionner que l'homme de Neandertal possède un caractère dérivé absent chez Homo erectus et Homo sapiens : l'absence de fosse canine (voir bibliographie). Cet argument étaye l'idée d'un rameau ayant évolué indépendamment d'Homo sapiens.
2. Homo sapiens sapiens
- Datés de 100 000 ans à l’actuel, de grande taille (1,7m), d'un grand volume cérébral (de 1100 à 2000 cm3, avec une moyenne de 1500 cm3), bipèdes permanents, maîtrisent le feu, ont un activité industrielle (des outils diversifiés en matériaux variés) et artistique peut-être depuis 50 000 ans (- La grotte Chauvet 32 000 ans -, peintures murales, sculptures, objets décoratifs), respectent des rites (dont les rites funéraires), sont présents en Eurasie.
E. L’hominisation : le critère majeur est le développement du cerveau
Document complémentaire : Comparaison des boîtes crâniennes et des moulages endocrâniens.![]()
F. Deux modèles pour l'émergence de l’Homme moderne
1. Modèle du « remplacement » s’appuyant sur l'étude de l’ADN mitochondrial
- Le premier modèle, dit du « remplacement » (ou encore « Out of Africa ») admet une origine africaine unique à l'homme moderne. A partir de ce berceau, l'homme moderne aurait ensuite essaimé dans le reste du monde et aurait remplacé les populations d'hommes archaïques pouvant encore y exister.
- Les arguments en faveur du premier modèle sont essentiellement basés sur la comparaison des séquences d'ADN mitochondrial dans différentes populations humaines. La population présentant la plus grande diversité génétique de cet ADN est africaine. Si on admet que le rythme de mutations est sensiblement constant, c'est donc cette population qui occupe le berceau d'où est issu l'homme moderne. Les auteurs ont pu suggérer que l'ancêtre commun des hommes modernes était une africaine vivant il y a 100 000 à 200 000 ans et surnommée « l'Eve mitochondriale ».
- L’ADN mitochondrial ou ADNmt est une petite molécule d'ADN contenue dans les mitochondries et représentant quelques dizaines de gènes, elle est transmise à la descendance par les femmes uniquement (seul, le noyau du spermatozoïde pénètre dans l'ovocyte). (Tiré du manuel Bordas)
2. Modèle de « l'origine multi-régionale »
- Le second modèle, dit de « l'origine multi-régionale », soutient que les différentes populations d'Homme moderne proviennent de lignées différentes de « précurseurs » (des proto-Cro-Magnons ?) qui auraient évolué chacun dans leur région à partir de populations d'Homo erectus.
- Les arguments en faveur du second modèle sont tirés de l'analyse des fossiles. Les partisans de ce modèle insistent sur la continuité de certains traits anatomiques depuis les hommes « archaïques » jusqu'aux hommes modernes en Europe et en Asie. (Tiré du manuel Bordas)